
Daniel Mota Rojas
Alexandra L. Whittaker
Ana C. Strappini
Marcelo Ghezzi
Adriana Domínguez Oliva
Julio Martínez Burnes
Fabio Napolitano
Agustín Orihuela
-
INTRODUCCIÓN
Los animales no humanos (tanto domésticos como aquellos de fauna silvestre) están expuestos a diversos estímulos nocivos que pueden culminar en la percepción del dolor (Mota-Rojas, 2013, 2014). La función primordial del dolor, el cual es una experiencia sensorial y emocional desagradable, es servir como un mecanismo de defensa para minimizar y prevenir el daño que un estímulo nocivo pueda generar en el organismo (Mota-Rojas et al., 2010; Fenton et al., 2015; Burrell, 2017; Martins, 2019; Lee y Neumeister, 2020; Raja et al., 2020). La neurobiología del dolor involucra cinco fases en las cuales un estímulo nocivo periférico se convierte en dolor consciente a través de fibras nerviosas, regiones espinales, y estructuras cerebrales (Basbaum et al., 2009; Milner y Doherty, 2015; Mota-Rojas y Cajiao, 2016; Lee y Neumeister, 2020). Conocer cómo es que un evento doloroso (p. ej., corte de cola, laminitis, mastitis, procedimiento quirúrgico, entre otros) genera reacciones fisiológicas, endocrinas y conductuales en los animales, es pieza clave en la medicina veterinaria debido a la obligación ética que tienen los profesionales de prevenir y tratar el dolor (Mota-Rojas et al.. 2010, 2018; Fenton et al., 2015; Hernández-Avalos et al., 2019; Mota-Rojas et al., 2024a,b,c,d).
La percepción del dolor comienza con la detección de estímulos potencialmente nocivos (químicos, mecánicos y térmicos) a través de neuronas especializadas: los nociceptores (Basbaum et al. 2009; Milner y Doherty, 2015; Lee y Neumeister, 2020). Los nociceptores conforman el primer paso para iniciar con la integración del dolor que culmina en la corteza somatosensorial, en donde el cerebro otorga los componentes sensoriales y emocionales del dolor consciente (Yam et al., 2018; Fregoso et al., 2019). Aunque el dolor cumple con un rol protector, cuando éste no se controla o se cronifica, se pueden presentar anormalidades como una sensibilización a las fibras nerviosas -tanto periféricas como centrales (Ossipov et al., 2010; Woolf, 2011; Gaynor y Muir, 2015).
Como resultado de la percepción del dolor, los animales no humanos desarrollan respuestas fisiológicas con el fin de mantener su homeostasis y preservar un adecuado nivel de bienestar (Gaynor y Muir, 2015; Saberi Afshar et al., 2017; da Silva et al., 2018; Mota-Rojas et al., 2021a,b,c,d) y es fundamental reconocerlo mediante técnicas novedosas como la termografía infrarroja, pupilometría, unidades de acción facial y escalas de evaluación miltimodales (Mota-Rojas y Orihuela, 2017a,b; Mota-Rojas, 2017a,b; Mota-Rojas y Ghezzi, 2020; Mota-Rojas et al., 2021e,f,g). El bienestar animal es parte integral de la formación profesional de los veterinarios a nivel mundial (Mota-Rojas et al., 2018) y debido a que el dolor tiene un efecto negativo en todos los dominios del bienestar (nutrición, ambiente, salud, comportamiento y estado mental), es de suma importancia que los clínicos comprendan los fundamentos neurobiológicos del dolor y su reconocimiento (Broom y Johnson, 2020; Mellor et al., 2020; Mota-Rojas et al., 2020, 2021). De acuerdo con el actual juramento veterinario, en el cual se menciona que los médicos veterinarios deben emplear sus conocimientos y habilidades a favor de la protección y cuidado de la salud y bienestar de los animales (FMVZ, 2024), así como el evitar el sufrimiento innecesario de los animales (Garcés, 2016; Mota-Rojas et al., 2018). La interacción humano animal positiva juega un rol clave para que los animales no perciban estados emocionales negativos como miedo o dolor y esto pueda afectar su nivel de productividad o su calidad de vida o muerte (Mota-Rojas et al., 2024c,d,e,f)
La implementación de estrategias efectivas de manejo del dolor no solo es un imperativo ético, sino que también promueve la recuperación, el comportamiento social y la interacción positiva con su entorno (Mota-Rojas et al., 2023a,b; Mota-Rojas et al., 2024e,f).
Figura 1. En medicina veterinaria se considera que la presencia o ausencia de dolor es un índice relacionado a la calidad de vida de los animales (Capner et al., 1999; Hugonnard et al., 2004; Mota-Rojas et al., 2023a,b). No obstante, aunque existen estas guías y escalas para evaluar el dolor, las bases neurobiológicas de éste suelen ser excluidas de los programas de estudio de los médicos veterinarios, por lo que el presente artículo busca ser una guía teórica para iniciarse en el estudio del dolor a través del análisis de los fundamentos neurobiológicos del dolor en animales no humanos.
-
NEUROBIOLOGÍA DEL DOLOR
El dolor se reconoce como una experiencia subjetiva, emocional y consciente que requiere un procesamiento neural en estructuras supraespinales (Basbaum et al., 2009; Milner y Doherty, 2015; Lee y Neumeister, 2020). La percepción consciente del dolor se lleva a cabo en el llamado “arco nociceptivo”, el cual es un conjunto de cinco fases que incluyen la transducción, transmisión, modulación, proyección y percepción (Kalueff et al., 2010; Gereau y Cavallone, 2013; Mota-Rojas, 2013, 2014; Bell, 2018; Yam et al., 2018; Martins, 2019; Hirose, 2022). La Figura 1 muestra estas cinco fases que culminan en los cambios fisiológicos, endocrinos y de comportamiento que se observan en los animales, etapas que serán discutidas en los siguientes subtemas (Hernández-Avalos et al., 2021a).
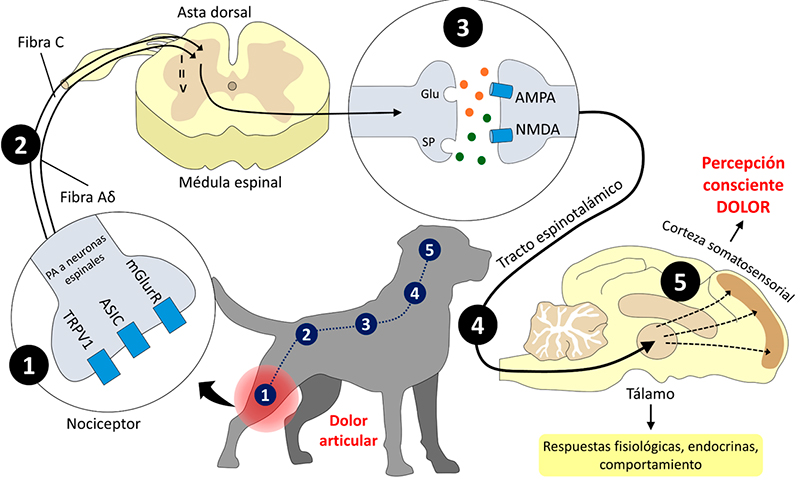
Figura 1. Esquematización del arco nociceptivo en un paciente canino. El procesamiento de un estímulo nocivo consta de cinco fases clave para la percepción consciente del dolor. (1) Transducción. Hace referencia a la transformación del estímulo nocivo térmico, químico o mecánico a una señal eléctrica para poder ser conducida a través de los axones de los nociceptores. Este cambio se realiza a través de los receptores localizados en los nociceptores periféricos. (2) Transmisión. La señal eléctrica es transmitida a lo largo de los axones de los nociceptores, particularmente de las fibras Aδ y C, para llegar al asta dorsal de la médula espinal. (3) Modulación. Este proceso se lleva a cabo en el asta dorsal de la médula espinal, en donde se estimulan o se inhiben las fibras nociceptivas. (4) Proyección. A través de las neuronas espinales, la información se conduce hacia el tálamo empleando vías ascendentes como la espinotalámica. (5) Percepción. Una vez que la información se encuentra en el tálamo, las conexiones de esta estructura con la corteza somatosensorial culminan en la percepción consciente del dolor. AMPA: α-amino-3-hidroxi-5-metilo-4-isoxazolpropiónico; ASIC: canales iónicos sensibles al ácido; DRG: ganglio dorsal de la médula espinal; GLU: glutamato; mGlurR: receptor metabotrópico de glutamato; NMDA: n-metil-D-asparatato; SP: sustancia P; TRPV1: receptor de potencial transitorio vaniloide 1.
-
TRANSDUCCIÓN
La primera etapa del arco nociceptivo requiere el reconocimiento de un daño tisular en regiones periféricas ubicadas en músculos, piel, huesos o vísceras (p. ej., proceso inflamatorio en la pezuña de un bovino con laminitis, herida quirúrgica por ovariohisterectomía en un paciente canino, o una herida por pelea con congénere en un felino doméstico). Este daño genera la activación de fibras nerviosas de terminales libres, o nociceptores, los cuales son neuronas primarias o de primer orden, de alto umbral y especializadas en reconocer estímulos nocivos de diferente naturaleza, sean mecánicos (mecanorreceptores), químicos (quimiorreceptores), térmicos (termorreceptores) y polimodales (Lemke, 2004; Vanderah, 2007; Zegarra- Piérola, 2007; Guevara, 2008; Mota-Rojas et al., 2010; Yam et al. 2018; Kendroud et al., 2022).
El término transducción se refiere a la transformación de dicha lesión tisular a una señal eléctrica para ser conducida hacia el sistema nervioso central (Basbaum et al., 2009). Esta señal eléctrica se genera por la activación de diversos receptores en los nociceptores (Guevara, 2008). La activación de los receptores genera un cambio de voltaje al interior de los nociceptores, lo que conlleva la despolarización y la generación de potenciales de acción que posteriormente son llevados las neuronas de segundo orden en la médula espinal (Lamont et al., 2000).
Uno de los principales receptores encargados de transducir información potencialmente nociva son los receptores de potencial transitorio o TRP, por sus siglas en inglés (transient potential receptor). Actualmente se reconoce que estos canales dependientes de voltaje se dividen en subfamilias y cada una tiene la capacidad de responder a estímulos de diferente naturaleza (Sneddon et al., 2014; Marwaha et al., 2016; Sneddon, 2018). En la subfamilia vanilloide (TRPV1, TRPV2, TRPV3, TRPV4), los receptores TRPV1 son los encargados de detectar temperaturas de calor nocivo por encima de los 42°C. De manera similar, la familia de melastatina (TRPM8) responde a temperaturas de frío nocivo con 10°C (Julius y Basbaum, 2001; Lamont, 2008; Swieboda et al., 2013; Lezama-García et al., 2022).
Además de los TRP, existen otros receptores que son activados directamente por las sustancias que se liberan después de un daño tisular (Rosenbaum y Simon, 2007; Wiese y Yaksh, 2009; Ji et al., 2014; Pinho-Ribeiro et al., 2017; Hernández-Avalos et al., 2021a). Por ejemplo, existen receptores que detectan cada uno de los mediadores inflamatorios post-lesión, incluyendo a los iones de hidrógeno (H+) y potasio (K++), moléculas de adenosina trifosfato (ATP), bradicinina, prostaglandina, leucotrienos, serotonina, histamina, sustancia P, tromboxanos, radicales libres, citocinas como interleucinas (IL) IL-1, IL-6 y factor de necrosis tumoral alfa (TNF-a), entre otros (Hassel y Dingledine, 2012; Oliveira et al., 2016). Estas sustancias provienen de la degranulación de mastocitos y de la activación de células inmunes para promover una respuesta inflamatoria y la recuperación del tejido (Carpenter y Dickenson, 2005; Binshtok et al., 2008; Gold y Gebhart, 2010).
3.1. Sensibilización periférica
Aunque la activación de los receptores ubicados en los nociceptores tiene como objetivo principal el transducir el estímulo nocivo a una señal eléctrica, cuando el estímulo es persistente se puede generar un efecto de sensibilización periférica (Latremoliere y Woolf, 2009; Mota-Rojas y Ghezzi, 2017a,b). La sensibilización periférica se refiere a una disminución en el umbral de activación de los nociceptores alrededor del sitio de lesión debido a la sobreestimulación de nociceptores (Willis Jr., 1985; Bolay y Moskowitz, 2002).
La presencia de los mediadores proinflamatorios provenientes de macrófagos, neutrófilos, mastocitos, eosinófilos, linfocitos, plaquetas, células endoteliales y fibroblastos altera la activación de los receptores que responden a dichas sustancias (Mota-Rojas, 2013, 2014; Vardeh y Naranjo, 2017). Por ejemplo, la liberación prolongada de prostaglandinas y bradicinina disminuyen el umbral de activación de los TRPV1 a 35°C (umbral de activación normal a 42°C) debido a la alteración de diversas vías de señalización entre las que destacan las fosfolipasas y las dependientes de monofosfato cíclico de adenosina (cAMP) (Ferreira et al., 2004; Wang et al., 2006; Velázquez et al., 2007). La entrada de iones y el cambio de polarización de la membrana de los nociceptores culmina en una propagación continua de potenciales de acción, incrementando la respuesta de estas fibras nerviosas (Figura 2) (Gangadharan y Kuner, 2013).
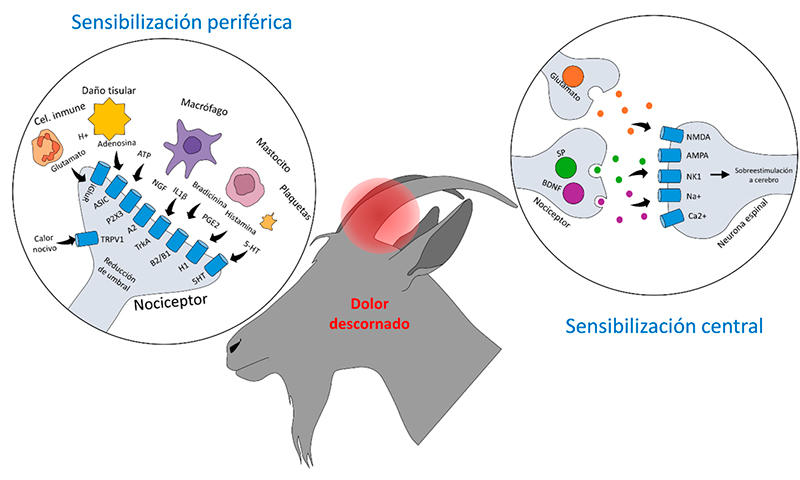
Figura 2. Mecanismos para la sensibilización periférica y central. La sensibilización central se produce por una sobreestimulación de los nociceptores periféricos. Durante este evento, los mediadores inflamatorios liberados después de un daño tisular interactúan directamente con los receptores ubicados en la membrana sináptica de los nociceptores. Esto genera cambios en la polarización de la membrana y la transmisión continua de potenciales de acción. Por otra parte, durante un evento de sensibilización central, las neuronas que participan son las espinales o de segundo orden, las cuales reaccionan a neurotransmisores excitatorios como el glutamato o la sustancia P, generando los mismos cambios de polarización membranal y una sobreexcitación neuronal que es conducida al cerebro. 5-HT: serotonina; A2: receptor de adenosina; AMPA: α-amino-3-hidroxi-5-metilo-4-isoxazolpropiónico; ASIC: canales iónicos sensibles al ácido; ATP: adenosín trifosfato; B1/B2: receptores de bradicinina; BDNF: factor neurotrófico derivado del cerebro; Ca2+: calcio; H+: hidrógeno; H1: receptor de histamina; IL: interleucina; Na+: sodio; NGF: factor de crecimiento neural; NK1: neurocinina 1; NMDA: n-metil-D-asparatato; P2X3: receptor prurinérgico; PGE2: prostaglandina E2; SP: sustancia P; TrkA: receptor de tropomiosina quinasa A; TRPV1: receptor de potencial transitorio vaniloide 1.
La sensibilización periférica también se acompaña de un efecto de hiperalgesia, el cual hace referencia a la reacción incrementada a un estímulo nocivo. La hiperalgesia se genera por la constante interacción de los mediadores proinflamatorios con sus respectivos receptores, y a la reducción del umbral de activación de dichos receptores (Mota-Rojas y Cajiao, 2016; Mota-Rojas y Ghezzi, 2017a). Además, se puede presentar la activación de los nociceptores silenciosos, los cuales no suelen participar en un evento de dolor agudo y controlado, pero que son cruciales para potenciar la sensación de dolor y aumentar la excitabilidad de las neuronas de primer orden (Julius y Basbaum, 2001; Zhang et al., 2005; Jensen y Finnerup, 2009; Schweizerhof et al., 2009; Gangadharan y Kuner, 2013). De igual forma, fibras nerviosas que se activan con estímulos de bajo umbral (las Aβ) y que normalmente no participan durante el arco nociceptivo, pueden contribuir en el procesamiento de señales nociceptivas durante un evento de sensibilización periférica (Ossipov et al., 2010).
-
TRANSMISIÓN
La transmisión de la señal eléctrica proveniente de la transducción de un estímulo nocivo se lleva a cabo en los axones de los nociceptores o neuronas de primer orden. Los principales nociceptores son las fibras nerviosas de terminales libres Aδ y C (Muir, 2010).
Las fibras tipo Ad se asocian con el “primer dolor”, caracterizado por una sensación aguda, rápida y punzante, debido a que son fibras mielinizadas (Lamont et al., 2000; Bell, 2018). La presencia de mielina les confiere una rápida velocidad de conducción de 12-30 m/seg. Son fibras que responden a estímulos nocivos mecánicos y térmicos (Zegarra-Piérola, 2007; Bosmans et al., 2009; Sneddon et al., 2014; Mertens et al., 2015; Mota-Rojas y Cajiao, 2016; Mota-Rojas y Ghezzi, 2017a). En contraste, las fibras amielínicas C procesan información química, mecánica y térmica, por lo cual se les conoce como nociceptores polimodales (Lamont et al., 2000; Bell, 2018). Debido a su falta de mielina, su velocidad de conducción es menor, de 0.5 a 2 m/seg, lo cual las hace las fibras responsables de transmitir los impulsos del “segundo dolor”, aquel que se reconoce por ser sordo, penetrante y visceral (Zegarra- Piérola, 2007; Bosmans et al., 2009; Swieboda et al., 2013; Sneddon et al., 2014; Mertens et al., 2015; Yam et al., 2018).
Los nociceptores también se clasifican de acuerdo con el tipo de estímulo que transmiten a las neuronas espinales o de segundo orden (Muir y Woolf, 2001; Vanderah, 2007; Leung, 2015). Los mecanorreceptores (tanto fibras Aδ como tipo C) procesan estímulos de tacto y presión; los termorreceptores responden a estímulos de calor y frío nocivo (desde 10°C a 45°C), mientras que los quimiorreceptores detectan alteraciones químicas como cambios en el pH o en la osmolaridad celular (Lamont, 2008; Swieboda et al., 2013).
Las fibras Aδ y C conducen la señal eléctrica desde la periferia hasta el asta dorsal de la médula espinal donde hacen sinapsis con las neuronas espinales o las de segundo orden (Gilron, 2000; Perena et al., 2000; Dubin y Patapoutian, 2010; Ossipov et al., 2010; Viet et al., 2014; Mertens et al., 2015). En el asta dorsal, estas neuronas hacen sinapsis con las neuronas espinales, también conocidas como de proyección o de segundo orden que se encuentran en la sustancia gris (Taylor y Robertson, 2004; Calvino y Grilo, 2006; Muir, 2010; Youn et al., 2017). A su vez, la sustancia gris de la médula espinal se divide en distintas partes conocidas como las láminas de Rexed. Las láminas I a la VI formar parte de la materia gris, la VII se ubica en la zona intermedia, las láminas VIII y IX están situadas en el cuerno ventral, y la lámina X circunda el canal central de la médula espinal.
Aunque existen diez láminas de Rexed, la información nociceptiva se procesa únicamente en las láminas I, II y V, en donde se localizan los nociceptores (Bosmans et al., 2009; Bourne et al., 2014; Sneddon et al., 2014). Cuando las fibras nociceptivas alcanzan las neuronas en el asta dorsal, éstas utilizan el tracto de Lissauer, que incluye neuronas propioespinales. Las neuronas Aδ realizan sinapsis en las láminas I, II y V, mientras que las neuronas C se vinculan con la lámina II que también tiene la capacidad de trasmitir información a las láminas I y V (Duke-Novakovski, 2017; Youn et al., 2017; Walters, 2018).
En la médula espinal también se encuentran otros tipos de neuronas, las propioespinales y las interneuronas. Tanto las neuronas de proyección, propioespinales y las interneuronas participan en el arco nociceptivo de manera distintiva. Por una parte, las neuronas de proyección transmiten la sensación desde la médula espinal hasta las estructuras del cerebro (Livingston, 2010; Bourne et al., 2014); las propioespinales participan generando los reflejos de retirada asociados a la nocicepción; y las interneuronas modulan los mecanismos de excitación e inhibición de la señal nociceptiva, el cual es la siguiente fase del arco nociceptivo (Bosmans et al., 2009).
-
MODULACIÓN
Esta etapa se refiere a los sistemas responsables de inhibir o potenciar las señales eléctricas que provienen de las interneuronas espinales (Ossipov et al., 2010; Moore, 2016). Esta fase del arco nociceptivo involucra neurotransmisores excitatorios tales como el ATP, la sustancia P y el glutamato. De igual manera, participan sustancia inhibitorias como el ácido gama aminobutírico (GABA), péptidos opioides endógenos (POE) y monoaminas (5-HT, NE, dopamina) (Mota-Rojas y Cajiao, 2016; Mota-Rojas y Ghezzi, 2017a; Yam et al., 2018; Puopolo, 2019). Estos compuestos se unen a los receptores localizados en la membrana sináptica de las interneuronas, principalmente a los receptores de N-metil-D-Aspartato (NMDA), ácido α-amino-3-hidroxi-5-metilo-4-isoxazolpropiónico (AMPA), kainato (KA), y metabotrópicos de glutamato (mGluR), cuya activación facilita la transmisión del impulso nociceptivo (Kwon et al., 2014; Kibaly et al., 2016; Harte et al., 2018; Malinowski, 2019).
Además de los neurotransmisores inhibitorios, los sistemas de modulación descendente tienen un papel fundamental para inhibir o controlar el dolor en animales. Estos sistemas requieren la integración y conexión neuronal de estructuras corticales prefrontales, el cíngulo anterior, corteza insular, amígdala y la sustancia gris periacueductal (PAG) (Kwon et al., 2014; Kibaly et al., 2016; Malinowski, 2019). A nivel cortical, en la PAG, durante el procesamiento de un estímulo doloroso se puede generar la liberación de POE, principalmente endorfinas (β-endorfinas), encefalinas (encefalin metionina, encefalin leucina), dinorfinas y nociceptinas (Kirkpatrick et al., 2015).
Los POE inhiben la señalización nociceptiva por su unión con receptores opioides μ, δ y κ, tanto a nivel central (generando hiperpolarización de la membrana) como periférico (evita al liberación de sustancias proinflamatorias) (Ossipov et al., 2004; Wang et al., 2006; Pérez, 2012; Flaherty, 2013; Heinricher, 2016; Bannister, 2019). No obstante, puede presentarse un efecto contrario conocido como “hiperalgesia inducida por opioides” cuando éstos son administrados a dosis repetidas, incrementando la excitabilidad de las neuronas e incrementando la trasmisión del estímulo doloroso (Kirkpatrick et al., 2015).
Otro de los sistemas descendentes de importancia es el serotoninérgico, ya que éste puede aumentar o disminuir la respuesta a un estímulo doloroso. En este sentido, la 5-HT puede ser antinociceptiva cuando se une a los receptores 5-HT1A, 5-HT1B, 5-HT1D, 5-HT7, propiciando una reducción o aumento en la secreción de glutamato, CGRP y SP (Ossipov et al., 2010; Bardoni, 2019). En contraste, su unión a los receptores 5-HT2A, 5-HT3 y TRPV1 lo hace pronociceptivo al disminuir la activación de dichos canales iónicos (Ossipov et al., 2010; Martins, 2019). En la PAG, amígdala y médula rostroventromedial (RVM) existen núcleos de neuronas serotoninérgicas del rafe magno (NRM) que se dirigen a la médula espinal mediante el funículo dorsolateral (Zhang et al., 2015). En esta región se encuentran núcleos de glicina y GABA que activan sistemas descendentes mediados por la 5-HT (Kirkpatrick et al., 2015; Lee y Neumeister, 2020).
De manera similar a la 5-HT y su ambivalencia en el arco nociceptivo, el sistema noadrenérgico localizado en el locus coeruleus es otro mecanismo que participa en la modulación del dolor, amplificando o disminuyendo la intensidad del mismo (Hayashida et al., 2010). La inhibición de un estímulo nociceptivo se genera con la activación de receptores espinales α2-adrenérgicos (α2A, α2B, α2C, α2D) (Yoon et al., 2021). En contraste, la amplificación del dolor se lleva a cabo cuando se activan los receptores α1-adrenérgicos (Fahy et al., 2021). El rol del sistema de modulación dopaminérgico (en el hipotálamo con proyecciones hacia la médula espinal) depende del receptor al que se una la dopamina, reconociendo que la activación de los receptores D1 y D5 generan hipersensibilización (Megat et al., 2018; Li et al., 2019; Puopolo, 2019). El sistema endocanabinoide (anandamida) es otro mecanismo de regulación que funciona mediante dos subtipos de receptores (CB1 y CB2) (Starowicz y Finn, 2017; Yam et al., 2018; Miranda-Cortés et al., 2023). Similar a la 5-HT, posee propiedades analgésicas y pro nociceptivas (Bouchet and Ingram, 2020; Hernández Ávalos, 2021b).
Finalmente, uno de los principales mecanismos de modulación es el conocido como “la teoría de la compuerta”, propuesto por Melzack y Wall (1965). Esta teoría hace referencia a la inhibición de un estímulo nocivo mediante la activación de fibras Aβ de la lámina II (Pereira y Lerner, 2017). Las fibras Aβ se reconocen como neuronas de un diámetro y una velocidad de conducción superior a la de los nociceptores (6–12 μm y 33-75 m/sec, respectivamente) (Kirkpatrick et al., 2015). Debido a ello, la activación de estas fibras a través de estímulos inocuos táctiles interrumpen la propagación de la señal eléctrica proveniente de las fibras A y C, promoviendo la analgesia (Bowditch, 2018). La teoría de la compuerta es la base teórica para técnicas como la electroestimulación nerviosa transcutánea, en la cual se activan las fibras Aβ para reducir el dolor e inflamación, sobre todo en eventos crónicos (Sato et al., 2020).
De esta manera, la modulación de la señal nociceptiva promueve la activación de neuronas inhibitorias o excitatorias en el asta dorsal de la médula espinal, señales que posteriormente son proyectadas a centros supraespinales para su posterior reconocimiento como dolor (Bagley y Ingram, 2020; Lee y Neumeister, 2020). Sin embargo, debido a que en la médula espinal se liberan una gran cantidad de neurotransmisores excitatatorios, cuando el dolor no es controlado se puede desarrollar sensibilización central.
5.1. Sensibilización central
La sensibilización central ocurre como consecuencia del incremento en la excitabilidad de las neuronas situadas en el asta dorsal de la médula espinal. La sobreestimulación de las neuronas espinales encargadas en procesar los estímulos nociceptivos genera una sobreproducción de neurotransmisores como el glutamato proveniente de fibras Ad y C (Mota-Rojas et al., 2010; Mota-Rojas, 2013, 2014; Vardeh y Naranjo, 2017). La presencia de glutamato y su unión a receptores NMDA, AMPA y KAI, disminuye el umbral de excitación de dichos receptores, generando la producción de potenciales de acción continuos que sensibilizan directamente las neuronas espinales (Sivilotti y Woolf, 1994; Baba et al., 2003; Ossipov et al., 2010).
Las células neuronales del asta dorsal que han sido sensibilizadas presentan un incremento en la actividad espontánea y en la respuesta a la estimulación, por lo que se facilita la transformación de estas neuronas a axones de proyección de señales nociceptivas (Sivilotti y Woolf, 1994; Ji et al., 2003; Latremoliere y Woolf, 2009; Woolf, 2011; Jensen y Finnerup, 2014). Este efecto se observa inclusive frente a estímulos que generalmente no causan dolor (p. ej., estimulación inocua táctil), como se observa en eventos de alodinia, contribuyendo al desarrollo del dolor crónico (Schaible et al., 2006; Bliddal y Danneskiold-Samsøe, 2007; Yunus, 2007).
-
PROYECCIÓN
La proyección se lleva a cabo a través de las neuronas de segundo orden. Estas son las encargadas de llevar la información nociceptiva a centros supraespinales como la médula, puente, mesencéfalo y, principalmente, el tálamo (Lorenz et al., 2011; McKune et al., 2017). Las neuronas emplean tractos nerviosos que surgen de las láminas de Rexed, entre los que se encuentran el espinotalámico, espinorreticular, espinomesencefálico, trigeminotalámico, espinoparabraquial y espinohipotalámico (DeLeo, 2006; Pedrajas y Molino, 2008; Wiese y Yaksh, 2009; Livingston, 2010).
De manera particular, el tracto espinotalámico es la principal vía de proyección del dolor. Ubicado en la lámina I y la IV, se compone de dos regiones, el neoespinotalámico (lateral) y paleoespinotalámico (medial), los cuales procesan diferente tipos de información. Por una parte, la porción lateral transmite el dolor rápido y otorga las características de localización, intensidad y duración del estímulo (Lee y Neumeister, 2020). Por el contrario, la porción medial se encarga del dolor lento o crónico, y también otorga el componente emocional al dolor (Zegarra, 2007; Bourne et al., 2014; Youn et al., 2017).
El sistema espinoreticular se compone de grupos de axones que se proyectan desde la médula espinal, en las láminas I, IV y VI, hasta el sistema reticular mesencefálico, la PAG, el núcleo parabraquial, y la amígdala. Este tracto participa activando mecanismos antinociceptivos a través de los sistemas descendentes de modulación (Perena et al., 2000; Bosmans et al., 2009; Bourne et al., 2014; Mertens et al., 2015; Youn et al., 2017). Por otra parte, el tracto espino-hipotalámico transmite estímulos nociceptivos y se encarga de generar las respuestas cardiovasculares y endocrinas autónomas como las alteraciones en la frecuencia cardiaca, presión arterial o frecuencia respiratoria de los animales (Lemke, 2004; Zegarra, 2007; Shilo y Pascoe, 2014). El tracto espino-parabraquial participa en el arco nociceptivo al otorgar el componente afectivo del dolor al hacer sinapsis en el tálamo y amígdala (Perena et al., 2000; Zegarra, 2007; Mertens et al., 2015).
Tanto el tracto espinotalámico como el espinorreticular son los principales canales de propagación de las señales dolorosas (DeLeo, 2006; Pedrajas y Molino, 2008; Wiese y Yaksh, 2009; Livingston, 2010). Las conexiones existen entre las neuronas espinales, el tálamo y otras estructuras como la amígdala o el hipotálamo son las que generan las respuestas fisiológicas de los animales frente al dolor (Mota-Rojas et al., 2020; Hernández-Avalos et al. 2021b; Sadler et al. 2022; Whittaker et al. 2023).
-
PERCEPCIÓN
Una vez que el estímulo nocivo llega al tálamo, las neuronas de tercer orden que parte de regiones talámicas hacia la corteza somatosensorial, es la vía por la cual un estímulo nocivo térmico, químico o táctil se transforma en la sensación consciente de dolor (Bannister, 2019; Fregoso et al., 2019). La corteza somatosensorial es la región que otorga los componentes sensorias al dolor (Cooley, 2015; Huang et al., 2020), mientras que el córtex del cíngulo anterior modula la parte afectiva y motivacional (Xiao et al., 2019). Algunos autores refieren que las conexiones entre la corteza somatosensorial y otras regiones cerebrales comprenden la posible sexta fase de la nocicepción, la llamada integración (Yam et al., 2018). La integración se refiere a la generación de las respuestas afectivas, de comportamiento y autónomas en respuesta a los componentes sensorial-discriminativo, afectivo-emocional, y evaluativo-cognitivo del dolor (Dzikiti et al., 2003; van Loon et al., 2010; Gorlin et al., 2016; Ellison, 2017; Kata et al., 2017; Lee y Neumeister, 2020; Mota-Rojas et al., 2021h; Grandin et al., 2023).
-
IMPORTANCIA DEL ARCO NOCICEPTIVO EN EL RECONOCIMIENTO CLÍNICO DEL DOLOR EN ANIMALES
El entender la neurobiología del dolor en medicina veterinaria no sólo sirve para reconocer que un estímulo doloroso necesita ser procesado por fibras periféricas y estructuras centrales, sino que en cada fase participan fibras nerviosas y regiones distintas del sistema nervioso central, lo cual es el fundamento teórico para el manejo del dolor.
Figura 3. Los médicos veterinarios tienen como obligación ética y profesional cuidar la salud de los animales (domésticos y de fauna silvestre). Por ello, el estudio del dolor y la inclusión de las bases teóricas ligadas a éste debe ser considerada en los programas de estudio como pieza clave para evitar el sufrimiento en animales y preservar su bienestar (Mota-Rojas et al., 2018).
Actualmente, el diseño de protocolos analgésicos multimodales considera las propiedades de cada fármaco y su acción en cada una de las etapas del arco nociceptivo (Slingsby, 2008; Cicirelli et al., 2022). Por ejemplo, durante las dos primera etapas (transducción y transmisión), las cuales se llevan a cabo en los nociceptores periféricos, el uso de bloqueos regionales con fármacos como lidocaína o bupivacaína, interrumpen la propagación de la eléctrica al actuar en los receptores que se activan con el dolor (Wolfe, 2018; Abendschön et al., 2020). A nivel supraespinal, el uso de inhibidores de la recaptación de 5-HT y NE, como el tramadol y el tapentadol, activan las vías descendentes serotoninérgicas y adrenérgicas, teniendo un efecto antinociceptivo en los animales (Kirkpatrick et al., 2015; Domínguez-Oliva et al., 2021). Asimismo, la comprensión de los eventos de sensibilización periférica y central, los mediadores inflamatorios que participan, y la respuesta de ciertos receptores ayuda a mejorar el tratamiento del dolor en pacientes crónicos (Gaynor y Muir, 2014; Mathews, 2018).

Figura 4. Comprender la neurobiología del dolor es clave para un correcto reconocimiento y manejo del dolor, así como para prevenir las consecuencias de un dolor no tratado (p. ej., sensibilización periférica o central y cronificación del dolor). Preservar el bienestar de los animales implica promover una buena salud y prevenir el efecto negativo que el dolor genera en el estado mental de las especies. Por ello, la habilidad para identificar y manejar el dolor de forma eficaz es prioridad para los profesionales en medicina veterinaria que buscan mejorar el bienestar y calidad de vida de los animales.
-
CONCLUSIONES
El estudio del dolor requiere conocer la neurobiología del mismo y las fases por las cuales un estímulo potencialmente doloroso se transforma en la experiencia consciente del dolor. La transducción, transmisión, modulación, proyección y percepción son etapas del arco nociceptivo en el que nociceptores periféricos responden a estímulos nocivos químico, térmicos o mecánicos, y esta información es proyectada hasta el tálamo y la corteza somatosensorial. La cascada de cambios fisiológicos, endocrinos y de comportamiento que se observan en los animales cuando perciben dolor es el resultado de la integración de dichas etapas.
Comprender la neurobiología del dolor es clave para un correcto reconocimiento y manejo del dolor, así como para prevenir las consecuencias de un dolor no tratado (p. ej., sensibilización periférica o central y cronificación del dolor). Preservar el bienestar de los animales implica promover una buena salud y prevenir el efecto negativo que el dolor genera en el estado mental de las especies. Por ello, la habilidad para identificar y manejar el dolor de forma eficaz es prioridad para los profesionales en medicina veterinaria que buscan mejorar el bienestar y calidad de vida de los animales.
Referencias
Abendschön, N., Senf, S., Deffner, P., Miller, R., Grott, A., Werner, J., Saller, A.M.; Reiser, J.; Weib, C.; Zablotski, Y.; Ficher, J.; Bergmann, S.; Erhard, M.H.; Baumgartner, C.; Ritzmann, M.; Zöls, S. Local Anesthesia in Piglets Undergoing Castration—A Comparative Study to Investigate the Analgesic Effects of Four Local Anesthetics Based on Defensive Behavior and Side Effects. Animals 2020, 10, 1752. https://doi.org/10.3390/ani10101752
Baba, H.; Ji, R.-R.; Kohno, T.; Moore, K.A.; Ataka, T.; Wakai, A.; Okamoto, M.; Woolf, C.J. Removal of GABAergic Inhibition Facilitates Polysynaptic A Fiber-Mediated Excitatory Transmission to the Superficial Spinal Dorsal Horn. Mol. Cell. Neurosci. 2003, 24, 818–830. https://doi.org/10.1016/S1044-7431(03)00236-7
Bagley, E.E.; Ingram, S.L. Endogenous Opioid Peptides in the Descending Pain Modulatory Circuit. Neuropharmacology 2020, 173, 108131. https://doi.org/10.1016/j.neuropharm.2020.108131
Bannister, K. Descending Pain Modulation: Influence and Impact. Curr. Opin. Physiol. 2019, 11, 62–66. https://doi.org/10.1016/j.cophys.2019.06.004
Bardoni, R. Serotonergic Modulation of Nociceptive Circuits in Spinal Cord Dorsal Horn. Curr. Neuropharmacol. 2019, 17, 1133–1145. https://doi.org/10.2174/1570159X17666191001123900
Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and Molecular Mechanisms of Pain. Cell 2009, 139, 267–284. https://doi.org/10.1016/j.cell.2009.09.028
Bell, A. The Neurobiology of Acute Pain. Vet. J. 2018, 237, 55–62.
Binshtok, A.M.; Wang, H.; Zimmermann, K.; Amaya, F.; Vardeh, D.; Shi, L.; Brenner, G.J.; Ji, R.-R.; Bean, B.P.; Woolf, C.J.; et al. Nociceptors Are Interleukin-1 Sensors. J. Neurosci. 2008, 28, 14062–14073. https://doi.org/10.1523/JNEUROSCI.3795-08.2008
Bliddal, H.; Danneskiold-Samsøe, B. Chronic Widespread Pain in the Spectrum of Rheumatological Diseases. Best Pract. Res. Clin. Rheumatol. 2007, 21, 391–402. https://doi.org/10.1016/j.berh.2007.03.005
Bolay, H.; Moskowitz, M.A. Mechanisms of Pain Modulation in Chronic Syndromes. Neurology 2002, 59, S2–S7. https://doi.org/10.1212/WNL.59.5_suppl_2.S2
Bosmans, T.; Doom, M.; Gasthuys, F.; Simoens, P.; Van Ham, L.; Polis, I. Perioperative Pain: Physiology and Pathophysiology. Vlaams Diergeneeskd. Tijdschr. 2009, 78, 302–313.
Bouchet, C.A.; Ingram, S.L. Cannabinoids in the Descending Pain Modulatory Circuit: Role in Inflammation. Pharmacol. Ther. 2020, 209, 107495. https://doi.org/10.1016/j.pharmthera.2020.107495
Bourne, S.; Machado, A.G.; Nagel, S.J. Basic Anatomy and Physiology of Pain Pathways. Neurosurg. Clin. N. Am. 2014, 25, 629–638. https://doi.org/10.1016/j.nec.2014.06.001
Bowditch, J. Where Do the Electrodes Go? The Clinical Use of TENS. Vet. Nurse 2018, 9, 546–549, doi:10.12968/vetn.2018.9.10.546.
Broom, D.M.; Johnson, K.G. Stress and Animal Welfare : Key Issues in the Biology of Humans and Other Animals. 2020.
Burrell, B.D. Comparative Biology of Pain: What Invertebrates Can Tell Us about How Nociception Works. J. Neurophysiol. 2017, 117, 1461–1473. https://doi.org/10.1152/jn.00600.2016
Calvino, B.; Grilo, R.M. Central Pain Control. Jt. Bone Spine 2006, 73, 10–16. https://doi.org/10.1016/j.jbspin.2004.11.006
Capner, A.; Lascelles, B.D.X.; Waterman-Pearson, A.E. Current British Veterinary Attitudes to Perioperative Analgesia for Dogs. Vet. Rec. 1999, 145, 95–99. https://doi.org/10.1136/vr.145.4.95
Carpenter, K.; Dickenson, A. Peripheral and Central Sensitization. In Core Topics in Pain; Cambridge University Press, 2005; pp. 29–36.
Cicirelli, V.; Burgio, M.; Lacalandra, G.M., Aiudi, G.G. Local and Regional Anaesthetic Techniques in Canine Ovariectomy: A Review of the Literature and Technique Description. Animals 2020, 12, 1920. https://doi.org/10.3390/ani12151920
Cooley, K. Physiology of Pain. In Pain Management for Veterinary Technicians and Nurses; Goldberg, M.., Shaffran, N., Eds.; WWiley Blackwell: Iowa, USA, 2015; pp. 30–41.
da Silva, M.L.; Maximino, C.; Siqueira-Silva, D.H. Nocifensive Behavior as Evidence for Sentient Pain in Fish. Anim. Sentience 2018, 3, 1365. https://doi.org/10.51291/2377-7478.1365
DeLeo, J.A. Basic Science of Pain. J. Bone Jt. Surg. 2006, 88, 58–62. https://doi.org/10.2106/JBJS.E.01286
Domínguez-Oliva, A.; Casas-Alvarado, A.; Miranda-Cortés, A.E.; Hernández-Avalos, I. Clinical Pharmacology of Tramadol and Tapentadol, and Their Therapeutic Efficacy in Different Models of Acute and Chronic Pain in Dogs and Cats. J. Adv. Vet. Anim. Res. 2021, 8, 404. https://doi.org/10.5455/javar.2021.h529
Dubin, A.E.; Patapoutian, A. Nociceptors: The Sensors of the Pain Pathway. J. Clin. Invest. 2010, 120, 3760–3772. https://doi.org/10.1172/JCI42843
Duke-Novakovski, T. Basics of Monitoring Equipment. Can. Vet. J. = La Rev. Vet. Can. 2017, 58, 1200–1208.
Dzikiti, T.B.; Hellebrekers, L.J.; Dijk, P. Effects of Intravenous Lidocaine on Isoflurane Concentration, Physiological Parameters, Metabolic Parameters and Stress-Related Hormones in Horses Undergoing Surgery. J. Vet. Med. Ser. A 2003, 50, 190–195. https://doi.org/10.1046/j.1439-0442.2003.00523.x
Ellison, D.L. Physiology of Pain. Crit. Care Nurs. Clin. North Am. 2017, 29, 397–406. https://doi.org/10.1016/j.cnc.2017.08.001
Facultad de Medicina Veterinaria y Zootecnia (FMVZ). Juramento profesional. 2024. Disponible en https://fmvz.unam.mx/fmvz/principal/archivos/Juramento.pdf (Consultado el 9 de octubre, 2024).
Fahy, E.J.; Griffin, M.; Lavin, C.; Abbas, D.; Longaker, M.T.; Wan, D. The Adrenergic System in Plastic and Reconstructive Surgery. Ann. Plast. Surg. 2021, 87, e62–e70. https://doi.org/10.1097/SAP.0000000000002706
Fenton, B.W.; Shih, E.; Zolton, J. The Neurobiology of Pain Perception in Normal and Persistent Pain. Pain Manag. 2015, 5, 297–317. https://doi.org/10.2217/pmt.15.27
Ferreira, J.; Da Silva, G.L.; Calixto, J.B. Contribution of Vanilloid Receptors to the Overt Nociception Induced by B 2 Kinin Receptor Activation in Mice. Br. J. Pharmacol. 2004, 141, 787–794. https://doi.org/10.1038/sj.bjp.0705546
Flaherty, D. Understanding the Mechanisms behind Acute Pain in Dogs and Cats. Vet. Nurse 2013, 4, 82–87. https://doi.org/10.12968/vetn.2013.4.2.82
Fregoso, G.; Wang, A.; Tseng, K.; Wang, J. Transition from Acute Tto Chronic Pain: Evaluating Risk for Chronic Postsurgical Pain. Pain Physician 2019, 22, 479–488.
Gangadharan, V.; Kuner, R. Pain Hypersensitivity Mechanisms at a Glance. Dis. Model. Mech. 2013, 6, 889–895. https://doi.org/10.1242/dmm.011502
Garcés, G.L.F. Fundamentación para una promesa bioética de los científicos que experimentan con animales (Parte 1). Cienc. Soc. Hum. 2016, 16, 261-271.
Gaynor, J.S.; Muir, W.W. Handbook of Veterinary Pain Management; 3rd. ed.; Elsevier: St. Louis Missouri, USA, 2015; ISBN 9780323089357.
Gereau, R.W.; Cavallone, L.F. Mechanisms of Pain Transmission and Transduction. In Anesthetic Pharmacology; Evers, A.S., Maze, M., Kharasch, Eds.; Cambridge University Press: London UK, 2013; pp. 227–247.
Gilron, I. Neural Blockade in the Evaluation and Management of Chronic Pain: An Overview. Pain Res. Manag. 2000, 5, 93–100. https://doi.org/10.1155/2000/546792
Gold, M.S.; Gebhart, G.F. Nociceptor Sensitization in Pain Pathogenesis. Nat. Med. 2010, 16, 1248–1257. https://doi.org/10.1038/nm.2235
Gorlin, A.; Rosenfeld, D.; Ramakrishna, H. Intravenous Sub-Anesthetic Ketamine for Perioperative Analgesia. J. Anaesthesiol. Clin. Pharmacol. 2016, 32, 160. https://doi.org/10.4103/0970-9185.182085
Guevara, L.U. Fisiopatología y Terapéutica Del Dolor Perioperatoria. Rev. Mex. Anestesiol. 2008, 31, S231–S234.
Grandin, T.; Velarde, A.; Strappini, A.; Gerritzen, M.; Ghezzi, M.; Martínez-Burnes, J.; Hernández-Ávalos, I.; Domínguez-Oliva, A.; Casas-Alvarado, A.; Mota-Rojas, D. Slaughtering ofWater Buffalo (Bubalus bubalis) with and without Stunning: A Focus on the Neurobiology of Pain, Hyperalgesia, and Sensitization. Animals 2023, 13, 2406. https://doi.org/10.3390/ani13152406
Harte, S.E.; Harris, R.E.; Clauw, D.J. The Neurobiology of Central Sensitization. J. Appl. Biobehav. Res. 2018, 23. https://doi.org/10.1111/jabr.12137
Hassel, B.; Dingledine, R. Glutamate and Glutamate Receptors. In Basic neurochemistry; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.N., Eds.; Elsevier, 2012; pp. 342–366.
Hayashida, S.; Oka, T.; Mera, T.; Tsuji, S. Repeated Social Defeat Stress Induces Chronic Hyperthermia in Rats. Physiol. Behav. 2010, 101, 124–131. ttps://doi.org/10.1016/j.physbeh.2010.04.027
Heinricher, M.M. Pain Modulation and the Transition from Acute to Chronic Pain. In; 2016; pp. 105–115.
Hernández-Avalos, I.; Mota-Rojas, D.; Mora-Medina, P.; Martínez-Burnes, J.; Casas Alvarado, A.; Verduzco-Mendoza, A.; Lezama-García, K.; Olmos-Hernandez, A. Review of Different Methods Used for Clinical Recognition and Assessment of Pain in Dogs and Cats. Int. J. Vet. Sci. Med. 2019, 7, 43–54. https://doi.org/10.1080/23144599.2019.1680044
Hernández-Avalos, I.; Flores-Gasca, E.; Mota-Rojas, D.; Casas-Alvarado, A.; Miranda-Cortés, A.E.; Domínguez-Oliva, A. Neurobiology of Anesthetic-Surgical Stress and Induced Behavioral Changes in Dogs and Cats: A Review. Vet. World 2021a, 14, 393–404. https://doi.org/10.14202/vetworld.2021.393-404
Hernández-Avalos, I.; Mota-Rojas, D.; Mendoza-Flores, J.E.; Casas-Alvarado, A.; Flores-Padilla, K.; Miranda-Cortes, A.E.; Torres-Bernal, F.; Gómez-Prado, J.; Mora-Medina, P. Nociceptive Pain and Anxiety in Equines: Physiological and Behavioral Alterations. Vet. World 2021b, 14, 2984–2995. https://doi.org/10.14202/vetworld.2021.2984-2995
Hirose, M. Nociception during Surgery. In Features and Assessments of Pain, Anaesthesia, and Analgesia; Rajendram, R., Patel, V.B., Preedy, V.R., Martin, C.R., Eds.; Academic Press: London, UK, 2022; pp. 235–245.
Huang, J.; Zhang, Z.; Zamponi, G.W. Pain: Integration of Sensory and Affective Aspects of Pain. Curr. Biol. 2020, 30, 393–395. https://doi.org/10.1016/j.cub.2020.02.056
Hugonnard, M.; Leblond, A.; Keroack, S.; Cadoré, J.; Troncy, E. Attitudes and Concerns of French Veterinarians towards Pain and Analgesia in Dogs and Cats. Vet. Anaesth. Analg. 2004, 31, 154–163. https://doi.org/10.1111/j.1467-2987.2004.00175.x
Jensen, T.S.; Finnerup, N.B. Neuropathic Pain: Peripheral and Central Mechanisms. Eur. J. Pain Suppl. 2009, 3, 33–36. https://doi.org/10.1016/j.eujps.2009.07.012
Jensen, T.S.; Finnerup, N.B. Allodynia and Hyperalgesia in Neuropathic Pain: Clinical Manifestations and Mechanisms. Lancet Neurol. 2014, 13, 924–935. https://doi.org/10.1016/S1474-4422(14)70102-4
Ji, R.-R.; Kohno, T.; Moore, K.A.; Woolf, C.J. Central Sensitization and LTP: Do Pain and Memory Share Similar Mechanisms? Trends Neurosci. 2003, 26, 696–705. https://doi.org/10.1016/j.tins.2003.09.017
Ji, R.-R.; Xu, Z.-Z.; Gao, Y.-J. Emerging Targets in Neuroinflammation-Driven Chronic Pain. Nat. Rev. Drug Discov. 2014, 13, 533–548. https://doi.org/10.1038/nrd4334
Julius, D.; Basbaum, A.I. Molecular Mechanisms of Nociception. Nature 2001, 413, 203–210. https://doi.org/10.1038/35093019
Kalueff, A.V.; Olivier, J.D.A.; Nonkes, L.J.P.; Homberg, J.R. Conserved Role for the Serotonin Transporter Gene in Rat and Mouse Neurobehavioral Endophenotypes. Neurosci. Biobehav. Rev. 2010, 34, 373–386. https://doi.org/10.1016/j.neubiorev.2009.08.003
Kata, C.I.; Rowland, S.; Goldberg, M.E. Pain Recognition in Companion Species, Horses, and Livestock. In Pain Management for Veterinary Technicians and Nurses; John Wiley & Sons, Ltd: Chichester, UK, 2017; pp. 15–29.
Kendroud, S.; Fitzgerald, L.A.; Murray, I. V; Hanna, A. Physiology, Nociceptive Pathways; StatPearls Publishing: Florida, USA, 2022.
Kibaly, C.; Loh, H.H.; Law, P.-Y. A Mechanistic Approach to the Development of Gene Therapy for Chronic Pain. In; 2016; pp. 89–161.
Kirkpatrick, D.R.; McEntire, D.M.; Hambsch, Z.J.; Kerfeld, M.J.; Smith, T.A.; Reisbig, M.D.; Youngblood, C.F.; Agrawal, D.K. Therapeutic Basis of Clinical Pain Modulation. Clin. Transl. Sci. 2015, 8, 848–856. https://doi.org/10.1111/cts.12282
Kwon, M.; Altin, M.; Duenas, H.; Alev, L. The Role of Descending Inhibitory Pathways on Chronic Pain Modulation and Clinical Implications. Pain Pract. 2014, 14, 656–667. https://doi.org/10.1111/papr.12145
Lamont, L.A.; Tranquilli, W.J.; Grimm, K.A. Physiology of Pain. Vet. Clin. North Am. Small Anim. Pract. 2000, 30, 703–728. https://doi.org/10.1016/S0195-5616(08)70003-2
Lamont, L.A. Multimodal Pain Management in Veterinary Medicine: The Physiologic Basis of Pharmacologic Therapies. Vet. Clin. North Am. Small Anim. Pract. 2008, 38, 1173–1186. https://doi.org/10.1016/j.cvsm.2008.06.005
Latremoliere, A.; Woolf, C.J. Central Sensitization: A Generator of Pain Hypersensitivity by Central Neural Plasticity. J. Pain 2009, 10, 895–926. https://doi.org/10.1016/j.jpain.2009.06.012
Lee, G.I.; Neumeister, M.W. Pain. Clin. Plast. Surg. 2020, 47, 173–180. https://doi.org/10.1016/j.cps.2019.11.001
Lemke, K.A. Understanding the Pathophysiology of Perioperative Pain. Can. Vet. J. = La Rev. Vet. Can. 2004, 45, 405–413.
Leung, E. Physiology of Pain. In Pain Management and Palliative Care; Sackheim, K., Ed.; Springer New York: New York, NY, 2015; pp. 3–6.
Lezama-García, K.; Mota-Rojas, D.; Pereira, A.M.F.; Martínez-Burnes, J.; Ghezzi, M.; Domínguez, A.; Gómez, J.; de Mira Geraldo, A.; Lendez, P.; Hernández-Avalos, I.; Falcón, I.; Olmos-Hernández, A.; Wang, D. Transient Receptor Potential (TRP) and Thermoregulation in Animals: Structural Biology and Neurophysiological Aspects. Animals 2022, 12, 106. https://doi.org/10.3390/ani12010106
Li, C.; Liu, S.; Lu, X.; Tao, F. Role of Descending Dopaminergic Pathways in Pain Modulation. Curr. Neuropharmacol. 2019, 17, 1176–1182. https://doi.org/10.2174/1570159X17666190430102531
Livingston, A. Pain and Analgesia in Domestic Animals. In Hand Exp Pharmacol; 2010; pp. 159–189.
Lorenz, M.D.; Coates, J.R.; Kent, M. Handbook of Veterinary Neurology; 5 th.; Elsevier: Missouri, USA, 2011.
Malinowski, M.N. Mediators of Pain and Pain Processing. In Deer’s Treatment of Pain; Springer International Publishing: Cham, 2019; pp. 61–73.
Martins, D. Serotonin and Nociception: From Nociceptive Transduction at the Periphery to Pain Modulation from the Brain. In The Serotonin System; Elsevier, 2019; pp. 203–224.
Marwaha, L.; Bansal, Y.; Singh, R.; Saroj, P.; Bhandari, R.; Kuhad, A. TRP Channels: Potential Drug Target for Neuropathic Pain. Inflammopharmacology 2016, 24, 305–317. https://doi.org/10.1007/s10787-016-0288-x
Mathews, K. Recognition, Assessment and Treatment of Pain in Dogs and Cat. Analg. Anesth. Ill or Inj. Dog Cat, 2018, 1, 81–111. https://doi.org/10.1002/9781119036500.ch7
McKune, C.M.; Murrell, J.C.; Nolan, A.M.; White, K.L.; Wright, B.D. Nociception and Pain. In Veterinary Anesthesia and Analgesia; John Wiley & Sons, Ltd: Chichester, UK, 2017; pp. 584–623.
Megat, S.; Shiers, S.; Moy, J.K.; Barragan-Iglesias, P.; Pradhan, G.; Seal, R.P.; Dussor, G.; Price, T.J. A Critical Role for Dopamine D5 Receptors in Pain Chronicity in Male Mice. J. Neurosci. 2018, 38, 379–397. https://doi.org/10.1523/JNEUROSCI.2110-17.2017
Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. https://doi.org/10.3390%2Fani10101870
Melzack, R.; Wall, P.D. Pain Mechanisms: A New Theory. Science (80-. ). 1965, 150, 971–979. https://doi.org/10.1126/science.150.3699.971
Mertens, P.; Blond, S.; David, R.; Rigoard, P. Anatomy, Physiology and Neurobiology of the Nociception: A Focus on Low Back Pain (Part A). Neurochirurgie 2015, 61, S22–S34. https://doi.org/10.1016/j.neuchi.2014.09.001
Milner, R.; Doherty, C. Pathophysiology of Pain in the Peripheral Nervous System. In Nerves and Nerve Injuries; Tubbs, R.S., Rizk, E., Shoja, M.M., Loukas, M., Barbaro, N., Spinner, R.J., Eds.; Academic Press: London UK, 2015; pp. 3–22.
Miranda-Cortés, A.; Mota-Rojas, D.; Crosignani-Outeda, N.; Casas-Alvarado, A.; Martínez-Burnes, J.; Olmos-Hernández, A.; Mora-Medina, P.; Verduzco-Mendoza, A.; Hernández-Ávalos, I. The Role of Cannabinoids in Pain Modulation in Companion Animals. Front. Vet. Sci. 2023, 9, 1050884. https://doi.org/10.3389/fvets.2022.1050884
Moore, S.A. Managing Neuropathic Pain in Dogs. Front. Vet. Sci. 2016, 3, 12. https://doi.org/10.3389/fvets.2016.00012
Mota-Rojas, D.; Guerrero-Legarrtea, I.; Trujillo, O.M. Bienestar animal y calidad de la carne. Valoración del dolor en animales que arriban al rastro. Primera edición. México. B.M. Editores, 2010. p. 361.
Mota-Rojas, D. Importancia de la etología en la evaluación del dolor en animales domésticos. Conferencia magistral. 2do.Curso de bioética y bienestar animal. Auditorio Jesús Vilchez. Universidad Autónoma Metropolitana. México. 2013.
Mota-Rojas, D. Monitorización del dolor en animales de granja sometidos a prácticas zootécnicas dolorosas. Conferencia magistral. 3er. curso de comportamiento, bioética y bienestar animal. Auditorio Jesús Vilchez. Universidad Autónoma Metropolitana. México. 2014.
Mota- Rojas, D.; Velarde, A.; Maris-Huertas, S.; Cajiao, M.N. Animal welfare, a global vision in Ibero-América. Bienestar Animal una visión global en Iberoamérica, 3rd. ed. Elsevier: Barcelona, Spain. 2016.
Mota-Rojas, D. y Cajiao, M.N. Neurobiología y reconocimiento del dolor y su impacto en la calidad de vida de animales domésticos y silvestres. Fundación Universitaria Agraria de Colombia. Uniagraria. Bogotá. Colombia. 2016.
Mota-Rojas, D.; Ghezzi, M. Reconocimiento del dolor y bienestar animal. 1ª. Jornada de Bienestar Animal. Facultad de Ciencias Veterinarias –Universidad Nacional del Centro de la Provincia de Buenos Aires Campus Universitario. UNCPBA. Buenos Aires, Argentina. 2017a.
Mota-Rojas, D.; Ghezzi, M. Reconocimiento del dolor durante la aplicación de métodos de aturdimiento y eutanasia. 1ª. Jornada de Bienestar Animal. Facultad de Ciencias Veterinarias –Universidad Nacional del Centro de la Provincia de Buenos Aires Campus Universitario. UNCPBA. Buenos Aires, Argentina. 2017b.
Mota-Rojas, D. Reconocimiento clínico del dolor en animales domésticos mediante las expresiones faciales. Conferencia magistral. 4to. curso de bioética y bienestar animal. Auditorio Universitario de Posgrado. Universidad Autónoma Metropolitana. México. 2017a.
Mota-Rojas, D. Análisis pupilar para la identificación del dolor en animales domésticos en proceso de parto. 5to. curso de bioética y bienestar animal. Auditorio Universitario de Posgrado. Universidad Autónoma Metropolitana. México. 2017b.
Mota-Rojas, D.; Orihuela, A.; Strappini-Asteggiano, A.; Nelly Cajiao-Pachón, M.; Agüera-Buendía, E.; Mora-Medina, P.; Ghezzi, M.; Alonso-Spilsbury, M. Teaching animal welfare in veterinary schools in Latin America. Int. J. Vet. Sci. Med. 2018, 6, 131–140. https://doi.org/10.1016/j.ijvsm.2018.07.003
Mota-Rojas, D.; Orihuela, A. Procesos neurofisiológicos y control autónomo de la pupila frente a estados emocionales negativos en animales domésticos. Seminarios de Neurofisiología del dolor y bienestar animal para estudiantes de Medicina Veterinaria. Conferencia Foro universitario 2019. Universidad Autónoma Metropolitana. Ciudad de Mexico. 2019a.
Mota-Rojas, D.; Orihuela A. Lenguaje corporal (orejas y cola), frente a estados emocionales negativos en animales domésticos. Seminarios de Neurofisiología del dolor y bienestar animal para estudiantes de Medicina Veterinaria. Conferencia Foro universitario 2019. Universidad Autónoma Metropolitana. Ciudad de Mexico. 2019b.
Mota-Rojas, D.; Ghezzi, M. Evaluación del dolor a través de la pupilometría. 8to. Curso de Bienestar Animal para estudiantes de Medicina Veterinaria. Conferencia en Foro universitario 2020. Universidad Autónoma Metropolitana. Ciudad de Mexico. 2020.
Mota-Rojas, D.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Hernández, E.; Martínez-Burnes, J.; Whittaker, A.L. The Utility of Grimace Scales for Practical Pain Assessment in Laboratory Animals. Animals 2020, 10, 1838. https://doi.org/10.3390/ani10101838
Mota-Rojas, D.; Miranda-Córtes, A.; Casas-Alvarado, A.; Mora-Medina, P.; Boscato, L.; Hernández-Ávalos, I. Neurobiology and modulation of stress-induced hyperthermia and fever in animals. Abanico Vet. 2021a, 11, 1–17.
Mota-Rojas, D.; Napolitano, F.; Strappini, A.; Orihuela, A.; Martínez-Burnes, J.; Hernández-Ávalos, I.; Mora-Medina, P.; Velarde, A. Quality of Death in Fighting Bulls during Bullfights: Neurobiology and Physiological Responses. Animals 2021b, 11, 2820. https://doi.org/10.3390/ani11102820
Mota-Rojas, D.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Lecona-Butrón, H.; Martínez-Burnes, J.; Mora-Medina, P.; Gómez-Prado, J.; Orihuela, A. Infrared thermal imaging associated with pain in laboratory animals. Exp. Anim. 2021c, 70, 1–12. https://doi.org/10.1538/expanim.20-0052
Mota-Rojas, D.; Marcet-Rius, M.; Ogi, A.; Hernández-Ávalos, I.; Mariti, C.; Martínez-Burnes, J.; Mora-Medina, P.; Casas, A.; Domínguez, A.; Reyes, B.; et al. Current Advances in Assessment of Dog’s Emotions, Facial Expressions, and Their Use for Clinical Recognition of Pain. Animals 2021d, 11, 3334, doi:10.3390/ani11113334.
Mota-Rojas, D. Temas controvesiales en bienestar animal. El dolor durante la muerte. Especialización en Bienestar Animal y Etología. Fundación Universitaria Agraria de Colombia. Uniagraria. Bogotá. Colombia. 2021e.
Mota-Rojas, D. Temas controvesiales en bienestar animal: El uso de la termografía infrarroja y la pupilometría para el reconocimiento del dolor. Especialización en Bienestar Animal y Etología. Fundación Universitaria Agraria de Colombia. Uniagraria. Bogotá. Colombia. 2021f.
Mota-Rojas, D. Practicas zootécnicas dolorosas: aspectos neurobiológicos y etológicos. Especialización en Bienestar Animal y Etología. Fundación Universitaria Agraria de Colombia. Uniagraria. Bogotá. Colombia. 2021g.
Mota-Rojas, D.; Napolitano, F.; Strappini, A.; Orihuela, A.; Ghezzi, M.D.; Hernández-Ávalos, I.; Mora-Medina, P.; Whittaker, A.L. Pain at the Slaughterhouse in Ruminants with a Focus on the Neurobiology of Sensitisation. Animals 2021h, 11, 1085. https://doi.org/10.3390/ani11041085
Mota-Rojas, D.; Martínez-Burnes, J.; Casas-Alvarado, A.; Gómez-Prado, J.; Hernández-Ávalos, I.; Domínguez-Oliva, A.; Lezama-García, K.; Jacome-Romero, J.; Rodríguez-González, D.; Pereira, A.M.F. Clinical usefulness of infrared thermography to detect sick animals: frequent and current cases. CABI Rev. 2022, 1-8. https://doi.org/10.1079/cabireviews202217040
Mota-Rojas, D., Ghezzi, M.D., Hermández-Avalos, I., Álvarez-Macías, A., Domínguez-Oliva, A., Napolitano, F., Lendez, P.A., Orihuela, A. Respuesta conductual y neurofisiológica al dolor durante la mastitis: eficiencia productiva de rumiantes lecheros, así como su relación con la terapia analgésica, antiinflamatoria y antimicrobiana. Soc. Rur. Prod. Med. Amb. 2023a, 23, 72-98.
Mota-Rojas, D.; Domínguez-Oliva, A.; Martínez-Burnes, J.; Casas-Alvarado, A.; Hernández-Ávalos, I. Euthanasia and Pain in Canine Patients with Terminal and Chronic-Degenerative Diseases: Ethical and Legal Aspects. Animals 2023b, 13, 1265. https://doi.org/10.3390/ani13071265
Mota-Roja, D.; Ghezzi, M.; Orihuela, A.; Buenhombre, J.; Pereira, A.M.F.; Martínez-Burnes, J. La pupilometría como método de valoración del dolor en Medicina Veterinaria. Sección Especial: “Aprendamos Juntos de Bienestar Animal”. Editorial BM Editores. Agosto. México City. Mexico. 2024a. p. 1-12. https://bmeditores.mx/secciones-especiales/la-pupilometria-como-metodo-de-valoracion-del-dolor-en-medicina-veterinaria/
Mota-Rojas, D.; Orihuela, A.; Hernández, A.I.; Domínguez, A.; Casas, A.; Lendez, P.A.; Ghezzi, M. Beneficios y consecuencias del recorte de pico en la productividad: Aspectos anatómicos, fisiológicos y neurobiología del dolor. Soc. Rur. Prod. Med. Amb. 2024b, 23, 99-118.
https://sociedadesruralesojs.xoc.uam.mx/index.php/srpma/article/view/503
Mota-Rojas, D.; Orihuela, A.; Domínguez, O.A.; Lendez, P.A.; Ghezzi, M.; Napolitano, F. Uso de anillo de goma para la castración en bovinos: neurobiología del dolor, aspectos legales, éticos y etológicos. Soc. Rur. Prod. Med. Amb. 2024c, 24, 83-105. https://sociedadesruralesojs.xoc.uam.mx/index.php/srpma/article/view/514
Mota-Rojas, D.; Napolitano, F.; Álvarez. M.A.; Lezama, G.K. Domínguez, O. A.: Reyes, S.B.; Orihuela, A. El descornado en bovinos: neurobiología del dolor y repercusiones conductuales. Soc. Rur. Prod. Med. Amb. 2024d, 24, 107-126. https://sociedadesruralesojs.xoc.uam.mx/index.php/srpma/article/view/515
Mota-Rojas, D.; Whittaker, A.L.; Strappini, A.C.; Orihuela, A.; Domínguez, A.; Mora, P.; Álvarez, A.; Hernández, I.; Olmos, A.; Reyes, B.; Grandin, T. Human animal relationships in Bos indicus cattle breeds addressed from a Five Domains welfare framework. Front. Vet. Sci. 2024e, 11, 1456120. https://doi.org/10.3389/fvets.2024.1456120
Mota-Rojas, D.; Whittaker, A.L.; Domínguez-Oliva, A.; Strappini, A.C.; Álvarez-Macías, A.; Mora-Medina, P.; Ghezzi, M.; Lendez, P.; Lezama-García, K.; Grandin, T. Tactile, Auditory, and Visual Stimulation as Sensory Enrichment for Dairy Cattle. Animals 2024f, 14, 1265. https://doi.org/10.3390/ani14091265
Muir III, W.W.; Woolf, C.J. Mechanisms of Pain and Their Therapeutic Implications. J. Am. Vet. Med. Assoc. 2001, 219, 1346–1356. https://doi.org/10.2460/javma.2001.219.1346
Muir, W.W. Pain: Mechanisms and Management in Horses. Vet. Clin. North Am. Equine Pract. 2010, 26, 467–480. https://doi.org/10.1016/j.cveq.2010.07.008
Oliveira, J.J.O.; Andrade, P.C.S.; Panossia, C.C. Inflammatory Mediators of Neuropathic Pain. Rev. Dor. Sao Paulo 2016, 17, 35–42.
Ossipov, M.H.; Lai, J.; King, T.; Vanderah, T.W.; Malan, T.P.; Hruby, V.J.; Porreca, F. Antinociceptive and Nociceptive Actions of Opioids. J. Neurobiol. 2004, 61, 126–148. https://doi.org/10.1002/neu.20091
Ossipov, M.H.; Dussor, G.O.; Porreca, F. Central Modulation of Pain. J. Clin. Invest. 2010, 120, 3779–3787. https://doi.org/10.1172/JCI43766
Pedrajas, J.M.; Molino, A.M. Bases Neuromédicas Del Dolor. Clin. y Sal 2008, 19, 277–293.
Pereira, P.J.S.; Lerner, E.A. Gate Control Theory Springs a Leak. Neuron 2017, 93, 723–724. https://doi.org/10.1016/j.neuron.2017.02.016
Perena, M.J.; Perena, M.F.; Rodrigo- Royo, M.D.; Romera, E. Neuroantomía Del Dolor. Rev. Soc. Esp. Dolor 2000, 7, 5–10.
Pérez, G.Y. ¿Son Utiles Los Opioides y Adyuvantes En El Dolor Agudo? Revisión Cuba. Anestesiol. y Reanim. 2012, 11, 48–56.
Pinho-Ribeiro, F.A.; Verri, W.A.; Chiu, I.M. Nociceptor Sensory Neuron–Immune Interactions in Pain and Inflammation. Trends Immunol. 2017, 38, 5–19. https://doi.org/10.1016/j.it.2016.10.001
Puopolo, M. The Hypothalamic-Spinal Dopaminergic System: A Target for Pain Modulation. Neural Regen. Res. 2019, 14, 925. https://doi.org/10.4103/1673-5374.250567
Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The Revised International Association for the Study of Pain Definition of Pain: Concepts, Challenges, and Compromises. Pain 2020, 161, 1976–1982. https://doi.org/10.1097/j.pain.0000000000001939
Rosenbaum, T., and Simon, S. A. (2007). TRPV1 Receptors and Signal Transduction.
Saberi Afshar, F.; Shekarian, M.; Baniadam, A.; Avizeh, R.; Najafzadeh, hossein; Pourmehdi, M. Comparison of Different Tools for Pain Assessment Following Ovariohysterectomy in Bitches. Iran. J. Vet. Med. 2017, 11, 255–265. https://doi.org/10.22059/ijvm.2017.138815.1004701
Sadler, K.E.; Mogil, J.S.; Stucky, C.L. Innovations and Advances in Modelling and Measuring Pain in Animals. Nat. Rev. Neurosci. 2022, 23, 70–85. https://doi.org/10.1038/s41583-021-00536-7
Sato, K.L.; Sanada, L.S.; Silva, M.D. da; Okubo, R.; Sluka, K.A. Transcutaneous Electrical Nerve Stimulation, Acupuncture, and Spinal Cord Stimulation on Neuropathic, Inflammatory and, Non-Inflammatory Pain in Rat Models. Korean J. Pain 2020, 33, 121–130. https://doi.org/10.3344/kjp.2020.33.2.121
Schaible, H.; Schmelz, M.; Tegeder, I. Pathophysiology and Treatment of Pain in Joint Disease☆. Adv. Drug Deliv. Rev. 2006, 58, 323–342. https://doi.org/10.1016/j.addr.2006.01.011
Schweizerhof, M.; Stösser, S.; Kurejova, M.; Njoo, C.; Gangadharan, V.; Agarwal, N.; Schmelz, M.; Bali, K.K.; Michalski, C.W.; Brugger, S.; et al. Hematopoietic Colony–Stimulating Factors Mediate Tumor-Nerve Interactions and Bone Cancer Pain. Nat. Med. 2009, 15, 802–807. https://doi.org/10.1038/nm.1976
Shilo, Y.; Pascoe, P.J. Anatomy, Physiology and Pathophysiology of Pain. In Pain management in veterinary practice; Egger, M.C., Love, L., Doherty, T., Eds.; Wiley- Blackwell, 2014; pp. 9–27.
Sivilotti, L.; Woolf, C.J. The Contribution of GABAA and Glycine Receptors to Central Sensitization: Disinhibition and Touch-Evoked Allodynia in the Spinal Cord. J. Neurophysiol. 1994, 72, 169–179. https://doi.org/10.1152/jn.1994.72.1.169
Slingsby, L. (2008). Multimodal analgesia for postoperative pain relief. In Pract. 30, 208–212. doi: 10.1136/inpract.30.4.208.
Sneddon, L.U.; Elwood, R.W.; Adamo, S.A.; Leach, M.C. Defining and Assessing Animal Pain. Anim. Behav. 2014, 97, 201–212. https://doi.org/10.1016/j.anbehav.2014.09.007
Sneddon, L.U. Comparative Physiology of Nociception and Pain. Physiology 2018, 33, 63–73. https://doi.org/10.1152/physiol.00022.2017
Starowicz, K.; Finn, D.P. Cannabinoids and Pain: Sites and Mechanisms of Action. Adv. Pharmacol. 2017, 80, 437–475. https://doi.org/10.1016/bs.apha.2017.05.003
Swieboda, P.; Filip, R.; Prystupa, A.; Drozd, M. Assessment of Pain: Types, Mechanism and Treatment. Ann. Agric. Environ. Med. 2013, Spec no. 1, 2–7.
Taylor, P.M.; Robertson, S.A. Pain Management in Cats—Past, Present and Future. Part 1. The Cat Is Unique. J. Feline Med. Surg. 2004, 6, 313–320. https://doi.org/10.1016/j.jfms.2003.10.003
van Loon, J.P.A.M.; Back, W.; Hellebrekers, L.J.; van Weeren, P.R. Application of a Composite Pain Scale to Objectively Monitor Horses with Somatic and Visceral Pain under Hospital Conditions. J. Equine Vet. Sci. 2010, 30, 641–649. https://doi.org/10.1016/j.jevs.2010.09.011
Vanderah, T.W. Fisiopatología de Dolor. Med. Clin. N. Am. 2007, 91, 1–12.
Vardeh, D.; Naranjo, J.F. Peripheral and Central Sensitization. In Pain Medicine; Yong, R., Nguyen, M., Nelson, E., Urman, R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 15–17.
Velazquez, K.; Mohammad, H.; Sweitzer, S. Protein Kinase C in Pain: Involvement of Multiple Isoforms. Pharmacol. Res. 2007, 55, 578–589. https://doi.org/10.1016/j.phrs.2007.04.006
Viet, C.T.; Corby, P.M.; Akinwande, A.; Schmidt, B.L. Review of Preclinical Studies on Treatment of Mucositis and Associated Pain. J. Dent. Res. 2014, 93, 868–875. https://doi.org/10.1177/0022034514540174
Walters, E.T. Defining Pain and Painful Sentience in Animals. Anim. Sentience 2018, 3, 1360. https://doi.org/10.51291/2377-7478.1360
Wang, H.; Ehnert, C.; Brenner, G.J.; Woolf, C.J. Bradykinin and Peripheral Sensitization. Biol. Chem. 2006, 387, 11–14. https://doi.org/10.1515/BC.2006.003
Whittaker, A.L.; Muns, R.; Wang, D.; Martínez-Burnes, J.; Hernández-Ávalos, I.; Casas-Alvarado, A.; Domínguez-Oliva, A.; Mota-Rojas, D. Assessment of Pain and Inflammation in Domestic Animals Using Infrared Thermography: A Narrative Review. Animals 2023, 13, 2065. https://doi.org/10.3390/ani13132065
Wiese, A.J.; Yaksh, T.L. Nociception and Pain Mechanism. In Handbook of Veterinary Pain Management; Gaynor, J.S., Muir, W.W., Eds.; Elsevier: USA, 2009.
Willis Jr., W.D. Central Nervous System Mechanisms for Pain Modulation. Stereotact. Funct. Neurosurg. 1985, 48, 153–165. https://doi.org/10.1159/000101121
Wolfe, C. (2018). On the farm procedures, anesthesia and analgesia. Am. Assoc. Bov. Pract. Conf. Proc., 126–127. doi: 10.21423/aabppro20173292.
Woolf, C.J. Central Sensitization: Implications for the Diagnosis and Treatment of Pain. Pain 2011, 152, S2–S15. https://doi.org/10.1016/j.pain.2010.09.030
Xiao, Z.; Martinez, E.; Kulkarni, P.M.; Zhang, Q.; Hou, Q.; Rosenberg, D.; Talay, R.; Shalot, L.; Zhou, H.; Wang, J.; et al. Cortical Pain Processing in the Rat Anterior Cingulate Cortex and Primary Somatosensory Cortex. Front. Cell. Neurosci. 2019, 13, 165. https://doi.org/10.3389/fncel.2019.00165
Yam, M.; Loh, Y.; Tan, C.; Khadijah Adam, S.; Abdul Manan, N.; Basir, R. General Pathways of Pain Sensation and the Major Neurotransmitters Involved in Pain Regulation. Int. J. Mol. Sci. 2018, 19, 2164. https://doi.org/10.3390/ijms19082164
Yoon, S.-Y.; Roh, D.-H.; Yeo, J.-H.; Woo, J.; Han, S.H.; Kim, K.-S. Analgesic Efficacy of Α2 Adrenergic Receptor Agonists Depends on the Chronic State of Neuropathic Pain: Role of Regulator of G Protein Signaling 4. Neuroscience 2021, 455, 177–194. https://doi.org/10.1016/j.neuroscience.2020.12.021
Youn, D.H.; Kim, T.W.; Cho, H.J. Pain in Animals: Anatomy, Physiology, and Behaviors. J. Vet. Clin. 2017, 34, 347–352. https://doi.org/10.17555/jvc.2017.10.34.5.347
Yunus, M.B. Role of Central Sensitization in Symptoms beyond Muscle Pain, and the Evaluation of a Patient with Widespread Pain. Best Pract. Res. Clin. Rheumatol. 2007, 21, 481–497. https://doi.org/10.1016/j.berh.2007.03.006
Zegarra- Piérola, J.W. Bases Fisiopatológicas Del Dolor. Acta Medica Peru. 2007, 24, 35–38.
Zhang, X.; Huang, J.; McNaughton, P.A. NGF Rapidly Increases Membrane Expression of TRPV1 Heat-Gated Ion Channels. EMBO J. 2005, 24, 4211–4223. https://doi.org/10.1038/sj.emboj.7600893
Zhang, Y.; Zhao, S.; Rodriguez, E.; Takatoh, J.; Han, B.-X.; Zhou, X.; Wang, F. Identifying Local and Descending Inputs for Primary Sensory Neurons. J. Clin. Invest. 2015, 125, 3782–3794. https://doi.org/10.1172/JCI81156




 en la Finalización de Cerdos: Pequeños Cambios,Grandes Resultados.")




